
灵长目(学名:Primates):是哺乳纲的1个目,共14科约51属560余种。大脑发达;眼眶朝向前方,眶间距窄;手和脚的趾(指)分开,大拇指灵活,多数能与其他趾(指)对握。分为原猴亚目和猿猴亚目:原猴亚目颜面似狐;无颊囊和臀胼胝;前肢短于后肢,拇指与大趾发达,能与其他指(趾)相对;尾不能卷曲或缺如。猿猴亚目颜面似人;大都具颊囊和臀胼胝;前肢大都长于后肢,大趾有的退化;尾长、有的能卷曲,有的无尾。按区域分布或鼻孔构造。

大多为杂食性,选择食物和取食方法各异。每年繁殖1-2次,每胎1仔,少数可多到3仔。幼体生长较慢。高等种类性成熟的雌性有月经,雄性能在任何时间交配。
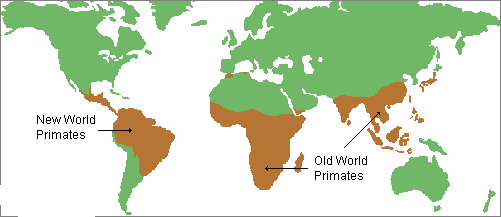
主要分布于亚洲、非洲和美洲温暖地带。是动物界最高等的类群。
- 灵长目
- Primates
- 动物界
- 脊索动物门
- 脊椎动物亚门
- 哺乳纲
- 真兽亚纲
- 灵长目
- 原猴亚目、简鼻亚目
- 14科
- 约51属
- 560余种
- Linnaeus, 175
物种学史
命名
灵长目与其他科属的翻译方式不太相同,灵长类的拉丁文名是Primates,这个命名源自二名法的创始人卡尔·冯·林奈,原意就是“首要,第一等”的意思 ,英语中的primary和这个词是同源词。而传到东方之后,日本人将拉丁文的Primates翻译为灵长,就是“众灵之长”,即动物进化的最高点。

灵长目中不同群体之间的关系一直到现代才比较清楚,因此一些常用的词可能会令人困惑,例如ape(猿)可能只是monkey(猴)的另一种称呼,但也可能只指没有尾巴,比较接近人的灵长目动物。
威尔弗里德·乐·格罗斯·克拉克爵士(Sir Wilfrid Le Gros Clark)是一位灵长类学家,他和其他灵长类学家发展了有关灵长目演化趋势的概念,并将未灭绝的灵长目依升序方式排序,最末端(即演化程度最高)的为人类。常用的名称像prosimian(原猴亚目)、monkeys(猴)、lesser apes(长臂猿)、great apes(人科)就是这种方法论下的产物。根据目前对灵长目演化的了解,这种分类是并系群,也就是该群中并不包含有最近共同祖先之所有后代。
化石
发现于北美普尔加托里山的普尔加托里猴是迄今为止全球最古老的灵长类动物化石。在灵长目中最早出现的是一些发现于欧洲和北美的近猴类化石。它们具爪而不具指甲。牙齿为三楔式低冠齿,比较一般化,但门齿增大,似平放的凿子。近猴类多发现于古新世地层。
自始新世开始狐猴类出现,早期的都归入已绝灭的兔猴科,它们的分布范围广,亚洲、北美、欧洲均曾发现。现在狐猴只分布于马达加斯加岛和科摩罗群岛,尚未发现可靠的化石。獭猴(又译瘦猴)现代只生存于东南亚和南亚、非洲撒哈拉以南的热带地区,化石发现于东非的中新世地层。眼镜猴类化石发现稍多。从始新世起发现于欧、亚、北美等地。近猴、狐猴、眼镜猴类常通称为原猴类或低等灵长类。
大约在2003年前后,考古人员在中国湖北省发现一具迄今最完整的早期灵长动物骨架化石,并与美国卡内基自然历史博物馆研究人员等国际同行展开合作研究,通过X射线技术对化石进行了高分辨率数字重建。研究显示,这种被命名为“阿喀琉斯基猴”的古猴生活于约5500万年前潮湿、炎热的湖边,是迄今发现的最早灵长目动物。这种古猴身长约7厘米,体重不超过30克,体积接近现代的小侏儒狐猴。它还具有修长的四肢、尖小的牙齿和大眼窝,证明它善于跳跃和利用四肢走动、以昆虫为食并拥有良好的视力。
研究人员认为,新发现为灵长目动物的早期进化研究提供了重要线索,这种古猴的存在进一步支持了灵长目动物最早出现在亚洲而不是非洲的论点。研究人员猜测,这种古猴可能在后来的进化中分化为眼镜猴和类人猿两支,而后者在数百万年的进化后分化出人类。
起源
在灵长目中最早出现的是一些发现于欧洲和北美的近猴类化石。多发现于古新世地层。自始新世开始狐猴类出现,早期的都归入已绝灭的兔猴科,它们的分布范围广,亚洲、北美、欧洲均曾发现,但是在非洲没有发现化石证据。到渐新世,已经出现了猿和猴,并且朝不同的方向进化。
据新华社巴黎2009年7月8日电,法国国家科研中心8日发表公报说,该机构与其他国家研究人员合作研究推测,在缅甸发现的类人猿化石可能说明这一物种起源于亚洲,而不是人们此前认为的非洲。此前科学家们一直认为,类人猿起源于非洲大陆,不过随着类人猿化石在缅甸的发现,上述理论受到质疑。
公报说,早在20年前,法国国家科研中心的古生物学家洛朗·马里沃就与多国考古学家合作,在中国、泰国、巴基斯坦和缅甸进行挖掘。2005年和2008年,他们在缅甸挖掘出土了多件近4000万年前的灵长目动物化石,其中一块下颌骨牙齿十分尖利,同时磨损也非常严重,说明这种动物习惯用牙咬开坚硬的果壳,吸取其中营养,而这种饮食习惯只在类人猿身上发现过,如南美洲的僧面猴。
2010年科学家在在利比亚发现的新类人猿化石显示,人类的“摇篮”可能在亚洲。以前有很多科学家相信,人类来自于非洲。然而新研究认为,人类的直系祖先可能在其他地方(最有可能是亚洲)发展壮大之后,才前往非洲,并统治了非洲大陆。科学家在利比亚撒哈拉沙漠发现了早期灵长类动物4个物种的化石,这些物种可能生活在3900万年前,但它们与同期或者更早在非洲生活的物种都不相同,这表明它们是在其他地方进化后才到达非洲的。其中的3个物种来自于灵长类家族中不同的分支,说明它们拥有同一个祖先,并且经过了很长时间才进化而成。在这段时期,也就是始新世时期,很少或者几乎没有已知的类人猿在非洲生存。
进化
现生的灵长类从低等到高等排列有树猴、狐猴和眼镜猴、猴、猿和包括我们人类在内。人是从猿发展分化来的。因此,人在动物界的位置也属于灵长类。
灵长类是有胎盘类中最高等的一类,它和有胎盘类的基干——食虫类的亲缘关系非常密切。在七千万年前白垩纪时出现一类原始食虫类,从原始食虫类适应辐射分化出许多分支,其中有一支即向灵长类方向发展。
灵长类与食虫类基本亲缘关系最好的证据是现在分布在中国西南、海南岛、东南亚的东方树鼩。它的身体大小如松鼠,有一长吻和长尾。脑较大,嗅区小,眼眶和颞区之间有一骨隔开。大拇指(趾)和大脚指(趾)与其他的趾有点分开。它的特点介于食虫类和灵长类之间,但更接近于灵长类。东方树鼩的食料中除食虫以外已有一部分是食“果实”。早期灵长类在进化的过程中的食料可能已由食“虫”逐渐改变食“果实”到食多种多样的“杂食”。食性的改变在灵长类的进化历程中是很重要的一步。
从某方面说灵长类的身体结构是最不特化的动物。不特化才有条件向更高级的进化方向发展。
灵长类的四肢很灵活,保留五趾的形式,大拇指(趾)与其他指(趾)分开,便于攀援和执握物体。灵长类的脑颅很大;眼也很大,具有双眼立体视觉的能力;眼眶与颞区之间有一骨隔开,在进步的灵长类中眼睛完全在被封闭的眼眶之中。这些特点在灵长类的进步过程中占有很重要的位置。
在新生代六千万年前古新世时,灵长类从原始树鲍向着不同方向发展,产生了原始狐猴和眼镜猴。到了五千万年始新世晚期时又从原始狐猴进化辐射产生了原始的猴和猿类。到1一2千万年前中新世又从当时古猿中分化出一支向人的方向发展,到大约三百万年前终于出现了最早能制造工具、有自觉能动性的人类。
现在的狐猴只限于生活在马达加斯加岛及其附近岛屿。但在古新世时的狐猴广泛分布于亚洲、欧洲和美洲有哈普鲁猴、更猴等。我国始新世的蓝田狐猴。现在的眼镜猴仅分布于马来亚、菲律宾、婆罗洲、西里伯等地;但在始新世时分布于我国的眼镜猴有黄河猴、秦岭卢氏猴等。
现生猿猴类可分阔鼻猴类和狭鼻猴类。阔鼻猴类也称新大陆猴类,主要分布于南美、中美以及墨西哥。狭鼻猴类(包括狭鼻猴、猿、人)也称旧大陆猴类,主要分布于旧大陆。猕猴分布于印度和我国华南一带,但在地史上猕猴化石在中国华北更新统地层中常有发现。
现生的猿类有分布于非洲的黑猩猩和大猩猩和分布于亚洲东南部的猩猩和长臂猿。这些猿类在外貌和面部表情上,在身体内部的结构上都与人相似。这说明人和猿有共同的祖先。最原始的猿类出现于三千多万年前的渐新世时期。到了一、二千万年前中新世时出现许多活跃的古猿。古猿中的一支后来下地向人的方向发展而成为人类。已知和人类关系最近的古猿有在巴基斯坦和印度发现的腊玛古猿。这种古猿犬齿较小,齿弓近抛物线形,颌骨从面部突出的程度较小;总之,形态与人最为接近中国云南开远小龙潭煤矿中发现的一千万年前的森林古猿牙齿化石及非洲肯尼亚一千多万年前地层中发现的肯尼亚古猿也可能属于从猿向人过渡的类型。古猿中其他几支继续在树上生活或很久以后再下来,后来发展成为猩猩、大猩猩和黑猩猩。长臂猿大概是从更早的渐新世的一种猿类发展来的。而从猿到人的转变过程更是一个漫长的过程。
形态特征
编辑灵长目的体型变化很大,东部大猩猩体重达200千克,最小的灵长类动物是西马达加斯加落叶林中最近被重新发现的小鼠狐猴,该种动物头部和身体的长度为6.2厘米,尾部长度为13.6厘米,平均重量为30.6克;或贝氏倭狐猴体重只有30克。灵长目动物的眼睛在脸的前面,有眉骨保护眼窝。视觉敏锐,类人猿有辨色能力,但是许多原猴没有。鼻子比其他的哺乳动物短,嗅觉退化。灵长目动物的脑相对于自身体重显得大而且重,并且很复杂。大部分灵长动物上下颌前方都有一排牙床。位于中间的叫门齿,之后是犬齿。犬齿之后是前臼齿,最后是臼齿。灵长目动物四肢会抓握,而且四肢都有5个趾头,接近人类特征。
灵长目的多数种类鼻子短,其嗅觉次于视觉、触觉和听觉,金丝猴属和豚尾叶猴属的鼻骨退化,形成上仰的鼻孔;长鼻猴属的鼻子大又长。多数种类的指和趾端均具扁甲,跖行性。长臂猿科和猩猩科的前肢比后肢长得多。猿类和人无尾,在有尾的种类中,其尾长差异很大,卷尾猴科大部分种类的尾巴具抓握功能。一些旧大陆猴(如狒狒)的脸部、臀部或胸部皮肤具鲜艳色彩,在繁殖期尤其显著。臀部有粗硬皮肤组成的硬块,称为臀胼胝。
多数种类在胸部或腋下有1对乳头,雄性的阴茎呈悬垂形,除人类和蜘蛛猴外多数具阴茎骨,雌体具双角子宫或单子宫。大多为杂食性,选择食物和取食方法各异。
原猴亚目的物种颜面似狐,眼较大,耳能转向,脑容量相对较小,额骨和下颌骨未愈合;无颊囊和臀胼胝;前肢短于后肢,拇指与大趾发达,能与其他指(趾)相对,五趾只能同时伸屈,不能单独活动,趾端有爪;尾不能卷曲或缺如。包括8科。
简鼻亚目的物种颜面似人;大都具颊囊和臀胼胝;前肢大都长于后肢,大趾有的退化;尾长、有的能卷曲,有的无尾。按区域分布或鼻孔构造,高等种类可分为阔鼻猴(新大陆猴)类和狭鼻猴(旧大陆猴)类。包括6科。

生活习性
生理特性
绝大多数灵长类都栖息在树上,这一点与大多数哺乳动物不同。在树上生活对于灵长类来说是不同寻常的。它们脚下没有土地可支撑,因此必须用四肢抓握树干。与此相适应,它们的四肢末端由早期哺乳动物的爪子逐渐转变为每个手指都能够单独活动的手;最后,拇指还能够与其余的各个手指对握。可想而知,这样的演化必定能够改善灵长类在树枝间活动所需要的抓握能力;更重要的是,拇指和食指指尖的对握可以形成环状,从而大大提高了手掌抓握物体的准确度。这一进化特征的出现不仅对早期灵长类搜寻昆虫等食物非常有利,而且对于后来灵长类可以用手灵巧地摆弄各种物体、直至最后能够制造和使用工具打下了基础。
与手部的灵巧活动相配合,灵长类发展了立体的视觉。双眼向前望着几乎是相同的目标,脑部就可以接受一对视觉的影象。经过了大脑的处理,影象就产生了深度、形象和距离的感觉。这样对灵长类在林间腾越行进是非常重要的。灵巧的手加上立体视觉,就使得灵长类能够从三维空间观察物体,用手把物体任意移动和拨弄。这都是灵长类充分掌握四周环境特质的先决条件,也是激发好奇心的原动力。
灵长类还发展出辨认颜色的能力,这很可能与它起源于大眼睛的早期夜行性哺乳动物有关。早期夜行性哺乳动物的大眼睛是为了在夜间增强对光线的敏感性的,但是当灵长类起源后,它们在白天越来越活跃,大眼睛内的视网膜就转变为能够接受不同的色彩。能够分辨颜色有助于灵长类分辨若干食物,特别是热带雨林茂密树枝上的果子。
这样,灵长类具备了一套独特的感觉器,能够把触觉、味觉、听觉、尤其是色觉和立体视觉感受到的各种信息输入脑中。脑接收外界的信息与日俱增,进而能够把各种信息分类排比,最终产生了智力的发展。这样的智慧,是任何其它动物都没有的,这也就是为什么我们把这类动物叫做“灵长类”的原因。
社会行为
人类与其他灵长目动物不仅体质特征很相似,而且社会行为也很相近。一般认为这主要是由于它们的大脑很发达,因此它们的行为方式也比其他动物复杂。
灵长目动物个体之间也有相互影响。最值得注意的是修饰对方的活动。他们彼此去除同伴身上的寄生虫,杂物等。此外,灵长目动物的未成年个体之间经常游戏。研究者认为这是在了解周围环境,或者是在训练力量,因为如想争得领导者的地位往往取决于力量。
灵长目动物能够发出声音,彼此进行信息交流。他们用一系列的喊叫声或者身体某部位的动作来传递信息。
灵长目动物一般都有一个“居住区”,就是通常活动的一定区域。居住区周围经常有群体成员进行巡逻。在居住区内,还按照群的大小以及食物的获取方式分成不同的小区域。居住区经常根据季节的改变而改变。有些种群会迁徙到经常去的区域,称为“核心区”。
灵长目动物的婴儿与其他动物不同,他们在刚出生时一直跟随母亲,生活完全由母亲来照顾,依偎在母亲的怀里。当然,不同的灵长类动物之间的母亲行为也不相同。许多灵长目种类的雄性对婴儿往往起到父母的责任,一起照料和保护婴儿。
非人灵长目动物的一个重要的社会行为是侵略行为。这种行为一般是通过姿势来恐吓对手,而不是通过激烈的搏斗来表现。搏斗一般只用于争夺在群体中的领袖地位时才用。灵长目动物具有很强的等级制度。雄性领袖具有优先挑选食物、选择雌性,并且有下级的成员服侍。但是在他受伤或生病时,他就有可能失去这种地位。
由于灵长目动物与人类有很近的亲缘关系,其生活方式也可能与最早的人类相仿。因此对研究人类的祖先有很大的帮助。
分布范围
非人类灵长类动物在其自然栖息地有限,主要是在新大陆的热带和亚热带地区。它们从来没有在澳大利亚和太平洋上的大多数岛屿生存。然而,从大约5千5百万年前开始,最早的灵长类动物也生活在北美和欧洲。
繁殖方式
每年繁殖1-2次,每胎1仔,少数可多到3仔。幼体生长比较缓慢 。性成熟的雌性有月经,雄性能在任何时间交配(低等猴类除外)。
下属分类
目 |
亚目 |
次目 |
超科 |
科 |
亚科 |
现存物种举例 |
|---|---|---|---|---|---|---|
灵长目 |
原猴亚目 |
狐猴次目 |
狐猴超科 |
狐猴 |
||
瘦猴超科 |
瘦猴 |
|||||
眼镜猴次目 |
眼镜猴 |
|||||
简鼻亚目 |
阔鼻猴次目 |
卷尾猴超科 |
卷尾猴 |
|||
狭鼻猴次目 |
人猿超科 |
长臂猿科 |
长臂猿 |
|||
猩猩科 |
猩猩 |
|||||
人科 |
南方古猿亚科 |
已灭绝 |
||||
人亚科 |
人 |
灵长目可以分为两个亚目:
原猴亚目
|
鼠狐猴科
Cheirogaleidae
|
鼠狐猴属 |
Cheirogaleus |
克氏鼠狐猴 |
Cheirogaleus crossleyi |
|
大鼠狐猴 |
Cheirogaleus major |
|
肥尾鼠狐猴 |
Cheirogaleus medius |
|
小铁灰鼠狐猴 |
Cheirogaleus minusculus |
|
希氏鼠狐猴 |
Cheirogaleus sibreei |
|
倭狐猴属 |
Microcebus |
|
